

sammendrag | innledning | problemstilling | metoder | resultater | diskusjon | litteratur
Åge Brabrand
Zoologisk Museum
Rapport nr. 179 – 1998 ISSN 0333-161x
Østensjøvannet i Oslo har vært sterkt forurenset i flere tiår på grunn av stor tilførsel og tilsig av avløpsvann. Det er i perioden 1990-92 gjennomført en betydelig rehabilitering av ledningsnettet, og vannkvaliteten i tilløpsbekkene er forbedret. På tross av dette er vannkvaliteten i selve Østensjøvannet fortsatt preget av høye konsentrasjoner av næringssalter, massive algeoppblomstringer og lavt siktedyp.
I Østensjøvannet har Oslo vann- og avløpsverk igangsatt en tiltaksplan i tre faser, der fase 1 har vært å innhente bakgrunnsmateriale, fase 2 har inneholdt praktiske undersøkelser og en fremtidig fase 3 med en tiltaksplan. De praktiske undersøkelser i fase 2 har bestått i å utrede alternative tiltak for å bedre vannkvaliteten i Østensjøvannet utover rehabilitering av avløpsledninger.
Et mulig tiltak i innsjøer for å bedre vannkvaliteten er å endre på fiskebestander. Laboratorium for ferskvannsøkologi og innlandsfiske ved Zoologisk museum, Universitetet i Oslo, har som en praktisk undersøkelse i fase 2 gjennomført en fiskeribiologisk undersøkelse av Østensjøvannet i 1998.
Den foreliggende undersøkelsen har hatt en todelt målsetting. Den skal dokumentere tilstanden i fiskesamfunnet, og resultatene skal kunne brukes for å vurdere fiskens betydning for vannkvaliteten. Det er tatt kontakt med Reidar Borgstrøm (NLH), Bjørn Mejdell Larsen (NINA), Olav Skulberg og Bjørn Faafeng (begge NIVA) og Jan Økland (UiO) ved diverse problemstillinger underveis, og alle takkes for hyggelige og konstruktive diskusjoner.
Oslo,
10.12.1998
Åge Brabrand
Brabrand, Å. 1998. Fiskesamfunnet i Østensjøvannet, Oslo kommune: Artssammensetning, dominans og vurdering av begrensende faktorer.Rapp. Lab. Ferskv.Økol. Innlandsfiske, Univ. i Oslo, 179, 19 s
Det er gjennomført en fiskeribiologisk undersøkelse i Østensjøvannet for å klarlegge fiskeribiologisk status i innsjøen med tanke på artssammensetning og dominansforhold på den ene siden, og en vurdering av hvilket grunnlag fiskebestanden har til å påvirke vannkvaliteten på den andre.
De fiskeartene som ble påvist var abbor, mort, gjedde og karuss. Av de totale fangstene dominerte abbor med 80% forekomst (basert på antall individer). Mort og karuss utgjorde henholdsvis 8,6% og 9,3%, mens gjedde utgjorde 2,1%. Basert på innsjøens generelle produksjon og de fiskearter som var tilstede var hovedinntrykket at fangstene både på Nordisk miljøgarnserie og på Jensens bunngarnserie var svært lave.
På Nordisk miljøgarnserie i juli ble det tatt usedvanlig lite fisk totalt sett, men noe abbor ble tatt rett utenfor vegetasjonsbeltet. Kun en mort ble tatt her, mens det ikke ble påvist fisk ute i de pelagiske områder, til tross for 20 timers fiske. Mengdeforholdet mellom fangster av abbor og mort på nordisk miljøgarnserie i slutten av juli 1998 var 70:1 basert på antall og 165:1 basert på vekt.
Med Jensens bunngarnserie ble det påvist abbor, gjedde, karuss og mort i de strandnære områder, mens det i pelagiske områder ble påvist mort, abbor og karuss. Totalfangstene var her noe større i antall, noe som skyldes større garnflate totalt sett, men fangst pr. innsatsenhet var svært lav. Mengdeforholdet mellom fangster av abbor og mort på Jensens bunngarnserie (supplert med 10 og 16 mm) i slutten av august var 3.5:1 basert på antall og 2.6:1 basert på vekt. Det ble tatt enkeltindivider av mort av til dels betydelig størrelse, og største mort veide 1075 gr., med en totallengde på 38,5 cm.
Mort viste ingen stagnasjon i vekst med økende alder. Dessuten manglet flere årsklasser, bl.a. mort med 1, 2 og 3 vekstsesonger. Rekrutteringen og samlet bestandsstørrelse av mort er derfor trolig meget lav.Abbor dominerte fangstene, men de totale fangstene av abbor var også betydelig lavere enn forventet, ut fra innsjøens produksjonsevne og at bestanden av mort er lav. Abbor var tilstede med alle årsklasser representert opp til 10 år. Aldersfordelingen viste jevn årlig rekruttering. Abbor viste heller ikke vekststagnasjon, men det var stor variasjon mellom vekstforløp hos individene i materialet.Karuss er lite fangbar på garn og denne arten er sannsynligvis underrepresentert i garnfangstene. Det ble funnet gamle individer (18-24 år) med lengde 35-37 cm. Det inntrer vekststagnasjon hos karuss eldre enn 3-5 år. Lengden er da 25-32 cm. Forekomst av damkaruss med annet vekstforløp kan ikke utelukkes. Av gjedde ble det fanget få individer, alle var hunner med jevn vekst uten vekststagnasjon.
Det konkluderes med at fangstene er merkelig små i forhold til innsjøens produktivitet (mort og abbor med < 1 fisk/100 m2 garnflate pr. time på bunngarn i littoralsonen). Dette sammen med at mort, abbor og gjedde ikke har vekststagnasjon viser at fisketettheten er lav i forhold til produksjonspotensial. Basert på innsjøens høye produksjonsevne og de fiskeartene som er tilstede burde fangstene av mort ligget i størrelsesorden 250-300 /100 m2garnflate pr. time, og med total dominans av mort. I slike bestander er mort vanligvis næringsbegrenset, og viser derved vekststagnasjon. Av abbor og mort har bare abbor jevn årlig rekruttering, og verken abbor eller mort viser stagnasjon i vekst, noe som nærmest bekrefter at disse bestandene ikke er næringsbegrenset.
Både mort og abbor synes derfor begrenset av en annen faktor enn næring. Fangstmengde, vekstforløp og alderssammensetning antyder at disse artene i årene forut for 1998 har vært begrenset av en eller annen faktor som begrenser rekrutteringen uten at veksten hos de gjenværende individer viser den vekststagnasjonen som er typisk for tette bestander.
Oksygensvinn som årsak til lave tettheter kan ikke utelukkes, men det er vanskelig å se at dette alene er den begrensende faktor, både fordi fiskedød uten påvist oksygenmangel ble påvist høsten 1995, og fordi målinger utført av OVA indikerer bedring av oksygenforholdene på ettervinteren på 1990-tallet sammenliknet med tidligere år.
Utover oksygensvinn som mulig dødelighetsfaktor bør giftproduserende blågrønnalger anses som en mulighet. Vannet er meget grunt og har svært liten gjennomstrømning. Fisk har ingen mulighet for å rømme ned under det algeproduserende sjikt eller til innløpsos i kritiske perioder. Når det periodevis utvikles meget høye forekomstene av blågrønnalger i Østensjøvannet vil dette kunne gi maksimal effekt av en eventuell algegift. NIVA har foretatt spredte observasjoner av forekomsten av blågrønnalger og toxinproduserende stammer, og biotester for akutt toksisitet og kjemiske analyser viser at blågrønnalger i Østensjøvannet produserer cyanotoksiner.
Den praktiske konsekvensen av en bedret vannkvalitet vil være at de begrensende faktorer vil endre karakter etterhvert som vannkvaliteten eventuelt bedres. Fra å være begrenset av en faktor som periodevis gir dødelighet (oksygensvinn, algegift) eller dårlig vekst, forventes det at bestandene etter hvert vil bli næringsbegrenset. Dette innebærer følgende endringer:
Økt biomasse og antall av fisk vil innebære at den relative betydningen av fiskens bidrag til eutrofiering vil øke. Dette vil være prosesser som må karakteriseres som langtidsendringer og som vil gå parallelt med en eventuell bedring av vannkvaliteten.
Forholdene i innsjøen må uansett karakteriseres som ustabile, også på lang sikt. Ustabiliteten mht. fisk henger sammen med at perioder med oksygensvinn opplagt kan inntreffe som en følge av høy produksjon og naturgitte forhold som lite vannvolum, liten tilførsel fra det lokale nedbørfelt og gjennomstrømning ved overføring fra Nøklevann.
Gjennomstrømning og tilførsel av vann er trolig en nøkkelfaktor for å stabilisere forholdene. Økt gjennomstrømning kan gi økt overlevelse av fisk i kritiske perioder. Drenering av den lokale overflateavrenningen har trolig redusert den naturlige vanngjennomstrømningen som i utgangspunktet har vært liten. Bidrag fra Nøklevann gjennom overføring vil derfor fortsatt ha stor positiv betydning.
Den foreliggende undersøkelse har som mandat å dokumentere fiskesamfunnets artssammensetning og mengdeforhold. Dette skal ses i sammenheng med vannkvaliteten, og resultatene kunne brukes senere for å vurdere fiskesamfunnets betydning for vannkvaliteten.
I Norsk institutt for vannforsknings (NIVA) ”Landsomfattende trofiundersøkelse av norske innsjøer” er det vist at Østensjøvannet er en av Norges mest forurensede innsjøer. Årsaken til dette er stor tilførsel av urenset avløpsvann over mange tiår, og i sommerperioden registreres høye konsentrasjoner av næringssalter, store algeoppblomstringer med dominans av blågrønne alger og et lavt siktedyp. På grunn av det rike fuglelivet er området fredet som naturreservat.
Vannkvaliteten i Østensjøvannet og tilløpsbekkene er overvåket av Oslo vann- og avløpsverk (OVA) siden slutten av 1970-tallet. Det vises til OVA-rapport (1998) for nærmere beskrivelse av vannkvalitet og utviklingen av denne i de senere år. I perioden 1990-92 ble det gjennomført en betydelig forbedring av ledningsnettet i nedlagsfeltet til Østensjøvannet og til tilløpsbekkene. Hovedkonklusjonen fra dette arbeidet er at dette førte til en betydelig forbedring av vannkvaliteten i bekkene, mens vannkvaliteten nærmest er uforandret i selve innsjøen.
I perioden 1979-94 var blågrønnalgeneAnabaena spp., Oscillatoria agardhii ogMicrocystis spp. de dominerende gruppene. NIVA har påvist giftproduserende stammer innen slektene Anabaena ogMicrocystis fra Østensjøvannet (Skulberg, pers. med). Massedød av fugl kan i flere tilfeller settes i sammenheng med nærvær og inntak av giftprodusende blågrønnalger (Skulberg, pers. med).
Det er dokumentert fiskedød ved en rekke anledninger i Østensjøvannet, og dette er ikke bare av ny dato. Allerede våren 1860 ble det funnet mengder av død ål langs breddene, og ålen regnes som utdødd i Østensjøvannet fra denne tiden, dels som en følge av kald vinter dette år, og at nyinnvandring av ålelarver fra sjøen til Østensjøvannet gjennom Loelva og Østensjøbekken ikke lenger var mulig pga. forurensning fra bebyggelse og spillvann fra industri (Brun, Høeg og Sæther 1965).
I februar 1903 og 1909 ble det observert masseutvandring av fisk fra Østensjøvannet og ut gjennom Østensjøbekken. Dette er beskrevet i detalj av Huitfeldt-Kaas (1909). Det var her en omfattende utvandring av fisk, vesentlig mort, litt abbor og noen få gjedde. Morten var av alle størrelser fra ”½ mark vægtige ned til et par tommer lange yngel”. Det ble fisket opp betydelige mengder mort i utløpselva, anslagsvis 20 ”tønder”, iht. en avisnotis. Vannprøve samlet inn personlig av Huitfeldt-Kaas under dagene (ca 20 februar 1909) med utvandring av fisk og analysert av hr. stadskjemiker Schmeld viste en oksygenkonsentrasjon på 2,15 cm3/L, og Huitfeldt-Kaas anså det for bevist at årsaken til utvandring skyldes oksygenmangel i selve Østensjøvannet. Dette til tross for at en vannkjemisk analyse i 1901 ga ”meget tilfredsstillende resultat” og at isen fra vannet var gjenstand for salg og bruk ved brennevinsbrennerier helt fram til 1921 (Sæther 1965). Huitfeldt-Kaas setter mangelen på oksygen i forbindelse med at vannet er grunt, har liten gjennomstrømning og at det både i 1903 og 1909 var snøfattige vintre med betydelig barfrost. Dette medførte at isen var tykk og at tilførsel av vann fra de få og små bekkene og fra Bogerudmyra disse vintrene var minimal pga. gjenfrysing.
I 1925 advarte imidlertid helserådet mot bruk av isen pga. kloakktilførsel pga. direkte utslipp av kloakk til vannet fra omkringliggende bebyggelse (Tveter 1925). Ørekyten ble regnet som utdødd i 1920-årene og dødelighet av mort ble observert ved flere anledninger i 1940-årene.
Fiskedød i Østensjøvannet er senere påvist ved flere anledninger, uten at det har vært mulig å kunne dokumentere årsaken til disse med samme grad av nøyaktighet som de i 1903 og 1909. Siste dokumenterte fiskedød var i november 1995, med omfattende død av årsunger av abbor som da kunne observeres under isen. Som ved de fleste andre tilfellene ble det ikke funnet noen entydig årsak, men verken akutt utslipp eller infeksjoner ble ved denne anledningen angitt som sannsynlig årsak (Brabrand 1995). Det ble også foretatt oksygenmålinger av Geir Solheim (OVA) umiddelbart etter at død fisk ble observert, og forholdene viste tilfredsstillende forhold for fisk, noe som for så vidt måtte forventes så tidlig etter islegging. Det er derfor viktig å merke seg at omfattende fiskedød ved denne anledning har skjedd pga. av en annen årsak enn oksygensvinn.
Periodevis lave oksygenkonsentrasjoner er imidlertid ofte observert i de bunnære vannmasser, både om vinteren og om sommeren (Sæther 1965, OVA 1995). Dette kan selvsagt gi direkte dødelighet, men gir også indirekte et betydelig mindre tilgjengelig vannvolum for fisk med sammentrengning og mulighet for betydelig predasjon på småfisk som resultat.
I en innsjø med fiskedød vil omfang (hyppighet i tid, mengden av død fisk) og selektivitet (bestemte arter eller størrelser) ha avgjørende betydning for hvilket fiskesamfunn vi observerer i innsjøen. Aldersfordelingen kan vise den relative styrken på de enkelte årsklasser og derved angi overlevelse de enkelte år. En dødelighetsfaktor som ikke virker hvert år, eller som har forskjellig omfang fra år til år vil gi ujevnt sterke årsklasser.
I tillegg vil en rekke biologiske forhold i den gjenværende bestand bli forandret, først og fremst næringskonkurranse, der liten bestand vil gi stor individuell vekst. Artssammen-setningen, artenes alderssammensetning og enkeltindividenes vekst kan derfor benyttes som en indikator for miljøforholdene i innsjøen, eller mer presist hvilke flaskehalser som fiskebestandene er påvirket av i tiden forut for observasjonen (flere år). Konkret vil hovedspørsmålet være om fiskebestanden er begrenset av en eller annen dødelighetsfaktor eller om bestandene ikke har slike belastninger.
Utover disse indikatorene i enkeltbestander vil mengdeforholdet mellom fiskeartene og dominans kunne angi begrensende faktorer.
Mengdeforhold og dominans vil også ha direkte innflytelse på den effekt fiskebestandene kan ha på vannkvaliteten gjennom nedbeiting av algespisende zooplankton og resirkulering av sedimentbundet fosfor. Dessuten vil en periode med god vannkvalitet eller en langtidsutvikling mot bedre vannkvalitet (mindre dødelighet) gi som resultat at tetthet og produksjon i bestandene (artsfordeling, størrelser) vil øke, og at innflytelsen av fisk på vannkvaliteten gjennom de nevnte mekanismene vil øke. Dette vil kunne forsinke den bedring av vannkvaliteten som ellers ville være forventet ut fra tiltak i nedbørfeltet, og for så vidt også av de vannkjemiske parametre fra selve innjøen.
Det er benyttet settegarn av typen nordisk miljøgarnserie og modifisert Jensenserie (supplert med 16 mm og 10 mm maskevidde). Begge seriene ble benyttet ved land rett utenfor vegetasjonsbeltet og midt ute i innsjøen, se Fig. 1.
Nordisk miljøgarnserie er et 30 m langt garn (høyde 1,5 m) bestående av følgende maskevidder innsydd i følgende rekkefølge: 43, 19.5, 6.25, 10, 55, 8, 12.5, 24, 15.5, 5, 35, 29 mm, der hver maskevidde er representert med 2,5 m. En slik lenke ble satt parallelt med land og en midt i innsjøen. Det ble fisket på dagtid og nattid.
Modifisert Jensenserie besto av enkeltgarn med dimensjon 25*1,5 m, og følgende maskevidder ble benyttet: 10, 16, 19.5, 22.5, 26, 29, 35, 39, 45, 52 mm. Et garn av hver maskevidde ble satt fra land og rett ut. Dessuten ble en lenke bestående av et garn av hver maskevidde satt midt i innsjøen. Jensenserien ble satt kl. 16-18, og trukket påfølgende morgen kl. 9-11.
Fangstmengde pr. innsats (CPUE) av de ulike artene for hver maskevidde ble beregnet. Fra samtlige fisk på både nordisk miljøgarnserie og på Jensenserie ble det tatt strukturer til aldersbestemmelse (karpefisk, abbor: gjellelokk og ørestein, fra gjedde: vingebein og ørestein).
Det ble fisket med landnot 13.10.1998, etter at en del av vannvegetasjonen var noe redusert i omfang. Nota hadde en totallengde på 50 m, høyde 6 m og maskevidde 5 mm i notposen. Nottrekk var vanskelig å gjennomføre, idet strandvegetasjon hindret selve trekkingen mot land. Dessuten var bunnen så bløt at nota raskt ble fylt med mudder. Tre teknisk gode trekk ble gjennomført i den nord-østre del av innsjøen.
Under notfisket ble det stedvis observert betydelig bestand av vanlig dammusling (Anodonta piscinalis). Denne ble ansett som utdødd fra 1950-årene til den dukket opp igjen med enkelte individer i 1993 (Spikkerud 1994).
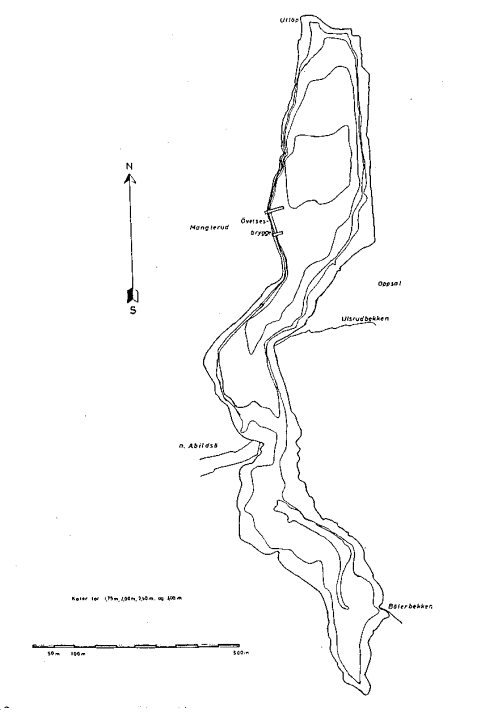
Fig. 1. Østensjøvannet i Oslo kommune, med angivelse av områder for notfiske, prøvefiske med bunngarn nær land og midt ute i innsjøen og transekter for kartlegging av vanlig dammusling. Dybdekart fra Sæther (1963).
Nottrekk:• . Transekt for undersøkelse av vanlig dammusling: A, B, C. Område for pelagisk fiske:
Område for fiske i strandsonen:
Dybdeutbredelse av liten dammusling ble derfor undersøkt ved dykking i uke 43 langs 4 transekter som strakte seg fra land og ca 100 m ut. Fire tetthetskategorier ble benyttet ved direkte observasjon av dykker:
Det ble tatt et tilfeldig utvalg av levende individer av vanlig dammusling for alderbestemmelse og lengdemåling (totallengde).
Fangstresultat på Nordisk miljøgarnserie er vist i Tabell 1 og Tabell 2 og for modifisert Jensen serie i Tabell 3 og Tabell 4. I samtlige tabeller er vist antall fisk fanget av de ulike arter fordelt på garnas maskevidde, og det er beregnet fangst pr. innsatsenhet (CPUE) angitt som antall fanget fisk pr. 100 m2 og time.
Det ble påvist gjedde, abbor, karuss og mort. Basert på innsjøens generelle produksjon og de fiskearter som var tilstede var hovedinntrykket at fangstene både på Nordisk miljøgarnserie og på Jensens bunngarnserie jevnt over var svært lave. På Nordisk miljøgarnserie i juli ble det tatt
Tabell 1. Fangstresultat (antall) og fangstmengde pr. 100 m2 garnflate og time (CPUE) for Nordisk miljøgarnserie i strandområder av Østensjøvannet 28-29.7.1998.
|
Maske-vidde |
Antall gjedde |
Antall mort |
Antall abbor |
Antall karuss |
CPUE gjedde |
CPUE mort |
CPUE abbor |
CPUE karuss |
|
5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
6.25 |
0 |
0 |
16 |
0 |
0 |
0 |
21 |
0 |
|
8 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
10 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
12.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
15.5 |
0 |
0 |
31 |
0 |
0 |
0 |
41 |
0 |
|
19.5 |
0 |
1 |
7 |
0 |
0 |
1,3 |
9 |
0 |
|
29 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
35 |
0 |
0 |
8 |
0 |
0 |
0 |
11 |
0 |
|
43 |
0 |
0 |
1 |
1 |
0 |
0 |
1,3 |
1,3 |
|
55 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Totalt |
0 |
1 |
70 |
1 |
0 |
0,11 |
7,8 |
0,11 |
Tabell 2. Fangstresultat (antall) og fangstmengde pr. 100 m2 garnflate og time (CPUE) for Nordisk miljøgarnserie i pelagiske områder av Østensjøvannet 28-29.8.1998.
|
Maske-vidde |
Antall gjedde |
Antall mort |
Antall abbor |
Antall karuss |
CPUE gjedde |
CPUE mort |
CPUE abbor |
CPUE karuss |
|
5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
6.25 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
8 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
10 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
12.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
15.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
19.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
24 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
29 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
35 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
43 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
55 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Totalt |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
Tabell 3. Fangstresultat og fangstmengde pr. 100 m2 garnflate og time (CPUE) for modifisert Jensen garnserie i strandområder av Østensjøvannet 26-27.8.1998.
|
Maske-vidde |
Antall gjedde |
Antall mort |
Antall abbor |
Antall karuss |
CPUE gjedde |
CPUE mort |
CPUE abbor |
CPUE karuss |
|
10 |
2 |
3 |
3 |
2 |
0,4 |
0,6 |
0,6 |
0,4 |
|
16 |
0 |
2 |
7 |
2 |
0 |
0,4 |
1,3 |
0,4 |
|
19.5 |
0 |
1 |
7 |
0 |
0 |
0,19 |
1,3 |
0 |
|
22.5 |
0 |
0 |
3 |
0 |
0 |
0 |
0,6 |
0 |
|
26 |
0 |
0 |
1 |
0 |
0 |
0 |
0,19 |
0 |
|
29 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
35 |
1 |
3 |
5 |
1 |
0,19 |
0,6 |
1,0 |
0,19 |
|
39 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
45 |
0 |
0 |
5 |
3 |
0 |
0 |
1,0 |
0,6 |
|
52 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Totalt |
3 |
9 |
31 |
8 |
0,06 |
0,17 |
0,59 |
0,15 |
Tabell 4. Fangstresultat og fangstmengde pr. 100 m2 garnflate og time (CPUE) for Jensen garnserie supplert med maskevidde 10 mm og 16 mm i pelagiske områder av Østensjøvannet 26-27.8.1998.
|
Maske-vidde |
Antall gjedde |
Antall mort |
Antall abbor |
Antall karuss |
CPUE gjedde |
CPUE mort |
CPUE abbor |
CPUE karuss |
|
10 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
16 |
0 |
0 |
6 |
0 |
0 |
0 |
1,1 |
0 |
|
19.5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
22.5 |
0 |
0 |
2 |
0 |
0 |
0 |
0,4 |
0 |
|
26 |
0 |
1 |
1 |
0 |
0 |
0,2 |
0,2 |
0,2 |
|
29 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
35 |
0 |
1 |
1 |
4 |
0 |
0,2 |
0,2 |
0,8 |
|
39 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
45 |
0 |
1 |
1 |
0 |
0 |
0,2 |
0,2 |
0 |
|
52 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Totalt |
0 |
3 |
11 |
4 |
0 |
0,06 |
0,21 |
0,08 |
usedvanlig lite fisk totalt sett, men noe abbor ble tatt rett utenfor vegetasjonsbeltet. Kun en mort ble tatt her, mens det ikke ble påvist fisk ute i de pelagiske områder, til tross for 20 timers fiske. Mengdeforholdet mellom fangster av abbor og mort på nordisk miljøgarnserie i slutten av juli 1998 var 70:1 basert på antall og 165:1 basert på vekt.
Med Jensens garnserie ble det til sammen påvist abbor, gjedde, karuss og mort i de strandnære områder, mens det i pelagiske områder ble påvist mort, abbor og karuss. Totalfangstene var her noe større i antall, noe som selvsagt skyldes større garnflate totalt sett, men fangst pr. innsatsenhet var svært lav. Mengdeforholdet mellom fangster av abbor og mort på Jensens modifiserte garnserie i slutten av august var 3,5:1 basert på antall og 2,6:1 basert på vekt. Det ble tatt enkeltindivider av mort med til dels betydelig størrelse, og største mort var 1075 gr. med en totallengde på 38,5 cm.
Det ble foretatt tre teknisk gode notdrag på dagtid. Det ble tatt abbor i lite antall i samtlige trekk, men ingen andre arter ble påvist med not. En abbor på 32 cm ble tatt, mens resten var årsunger i lengdeintervallet 6,4-7,2 cm.
For abbor viser aldersfordelingen at mange årsklasser er tilstede, med avtagende forekomst med økende alder (Fig. 2). Årsklassen 1998 (0+), er svakt representert, trolig fordi de ennå ikke er fullt ut fangbare på garn.
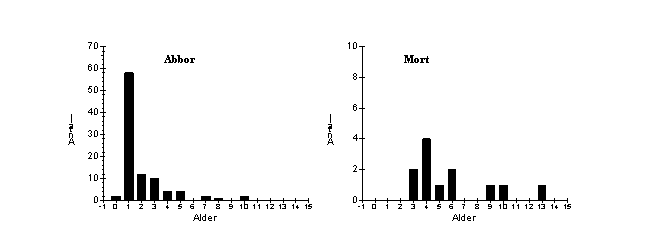
Fig. 2. Aldersfordeling av totalmaterialet av abbor og mort tatt under prøvefiske med bunngarn (nordisk miljøgarnserie og modifisert Jensen garnserie) i Østensjøvannet i juli og august 1998.
Aldersfordelingen for mort viste fravær av enkelte årsklasser (Fig. 2), men dette kan skyldes tilfeldigheter fordi totalfangsten av mort er lav. I materialet er det funnet mort opp til alder 13 år.
Begge kjønn hos abbor viser god vekst, med lengde etter første vekstsesong på 69,5 mm (SD=19,5) for hunner og 66,2 mm (SD=9,6) for hanner (Fig. 3). For hunner observeres en jevnt avtagende vekst, uten at dette kan karakteriseres som vekststagnasjon. For de tre individene i materialet som oppnår alder på 10 år påvises økt lengde fra alder 7 til 10 år.
Dette fremkommer tydeligere på individuelt vekstforløp (Fig. 5). Her fremkommer tydelig stor forskjell i individuell vekst, og ingen av individene har markert vekststagnasjon.
Mort viser jevn vekst frem til alder på 3-4 år for hunner og frem til 3 år for hanner (Fig. 3). Det individuelle vekstforløpet varierer svært mye, og for hunner av mort er det variasjon i lengden fra ca 12 cm til 30 cm ved 6 års alder (Fig. 5). Dette bidrar til at gjennomsnittlig vekst for aldersgruppene 5 og 6 åringer reduseres av enkelte individer som har svært lav vekst. Verken for individer med lav vekst eller god vekst er det vekststagnasjon. For de eldste individene er det fortsatt lengdevekst selv etter alder 7 år og frem til 13 år.
I Fig. 5 er vist individuell vekst for fire individer av karuss for å illustrere forskjell i vekstmønster. Tre individer har rask vekst frem til 3-5 års alder og viser deretter vekst-stagnasjon ved en lengde på ca 30 cm. Flere individer i materialet av karuss med lengde 35-37 cm viste en alder på 15-24 år, noe som antyder høy alder og vekststagnasjon for denne gruppen av karuss. Hvorvidt det finnes en gruppe karuss med et annet vekstmønster kan ikke angis med det foreliggende materialet.
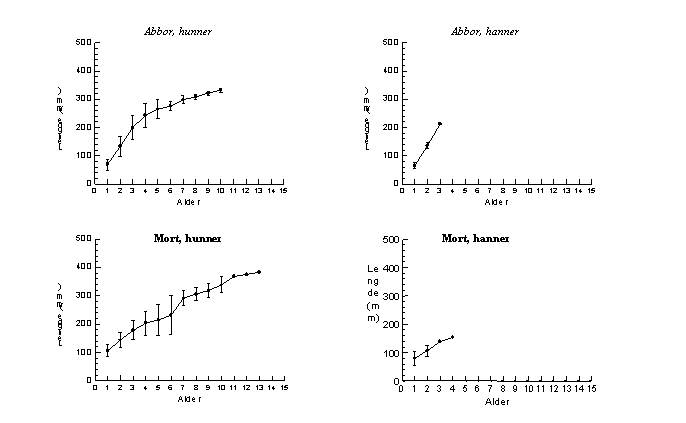
Fig. 3. Tilbakeberegnet vekst hos hunner og hanner av abbor (over) og av mort (under) tatt under prøvefiske med bunngarn (nordisk miljøgarnserie og modifisert Jensen garnserie) i Østensjøvannet i juli og august 1998.
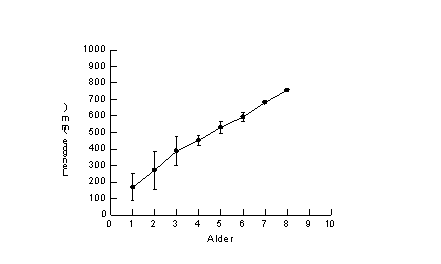
Fig. 4. Tilbakeberegnet vekst hos gjedde tatt under prøvefiske med bunngarn (nordisk miljøgarnserie og modifisert Jensen garnserie) i Østensjøvannet i juli og august 1998.
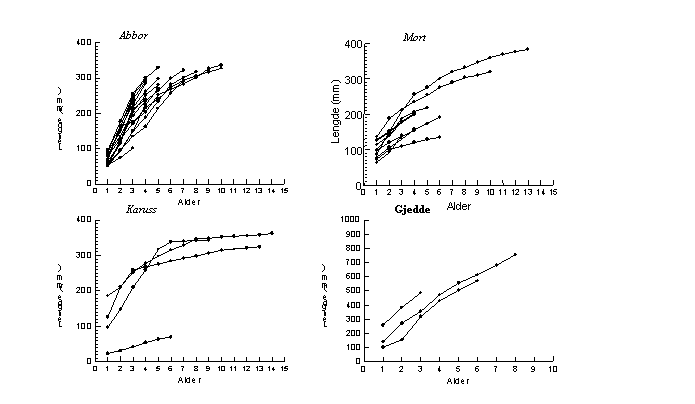
Fig. 5. Tilbakeberegnet vekst hos enkeltindivider av abbor, mort, karuss og gjedde tatt under prøvefiske med bunngarn (nordisk miljøgarnserie og modifisert Jensen garnserie) i Østensjøvannet i juli og august 1998.
Materialet av gjedde består kun av tre individer, alle hunner (samlet vekst vist i Fig. 4 og individuell vekst i Fig. 5). Disse viste jevnt god vekst og ingen tegn til vekststagnasjon frem til alder 8 år. Det var imidlertid stor variasjon mellom de tre individene i første 20 individer var de fleste aldersgrupper representert, men med dominans av individer med 5 vintersoner, dvs. årsklassen fra 1993.
De arter som i dag regnes som tilhørende i Østensjøvannet med regulære bestander er:
Karuss er ikke nevnt av Huitfeldt-Kaas (1918), og det har ikke vært mulig å dokumentere om denne arten er oversett av han, eller om den er innført på et seinere tidspunkt. Han nevner imidlertid ål, ørekyt og 9-pigget stingsild, men disse regnes som utryddet, ål allerede på 1860 – tallet på grunn av vandringshinder i Østensjøbekken, ørekyt og stingsild sannsynligvis knyttet til forurensning på 1940-tallet.
Huitfeldt-Kaas (1918) nevner forekomst av kreps i Loelva, men ikke spesielt i Østensjøvannet eller i Østensjøbekken. Kreps forekommer imidlertid i Nøklevann.
På grunnlag av prøvefiske i Østensjøvannet med bunngarn i strandområder og ute i vannmassene ble det i juli og august 1998 påvist abbor, mort, karuss og gjedde. Av de totale fangstene dominerte abbor med 80% forekomst. Mort og karuss utgjorde henholdsvis 8,6% og 9,3%, mens gjedde utgjorde 2,1%. Følgende forhold i resultatene poengteres:
Det konkluderes med at fangstene er merkelig små i forhold til innsjøens produktivitet og at dette reflekterer liten bestand for abbor, gjedde og spesielt for mort. Basert på innsjøens høye produksjonsevne og de fiskeartene som er tilstede skulle det ut fra dette alene forventes total dominans av mort, og at totalfangstene var betydelig større og i en helt annen størrelsesorden. I slike bestander er mort næringsbegrenset, og viser derved vekststagnasjon. Av abbor og mort har bare abbor jevn årlig rekruttering, og verken abbor eller mort viser stagnasjon i vekst, noe som nærmest bekrefter at disse bestandene ikke er næringsbegrenset.
Det er nærliggende å sammenligne fangstene med de som er rapportert fra Årungen. Denne innsjøen har lavere produktivitet, men har et fiskesamfunn bestående av mort, abbor, gjedde, ål og trolig karuss, dvs. forholdsvis likt det i Østensjøvannet. Årungen er imidlertid dypere og det er ikke rapportert om fiskedød. Fangstene ligger her på et helt annet nivå, 250-300 mort/100 m2 og time (Borgstrøm og Eie 1981), sammenliknet med Østensjøvannet som har < 1 fisk/100 m2 og time både for mort og abbor.
Både mort og abbor synes derfor begrenset av en annen faktor enn næring. Fangstmengde, vekstforløp og alderssammensetning antyder at disse artene i årene forut for 1998 har vært begrenset av en eller annen faktor som begrenser rekrutteringen uten at veksten hos de gjenværende individer viser den typiske vekststagnasjonen som er typisk for tette bestander. Derimot fortsetter veksten selv om individene er relativt gamle, men det er stor variasjon i selve forløpet mellom individene både hos mort og abbor. Hos karuss derimot antyder vekststagnasjon at bestanden er næringsbegrenset, og det er sannsynlig at bestandstettheten er større enn fangstene skulle tilsi.
En faktor som periodevis oksygensvinn skulle forventes å virke på bestandene i denne retningen. Mort er følsom for oksygensvinn, og er mer følsom enn abbor. I tillegg kan karuss overleve korte perioder under nærmest anaerobe forhold. Liten totalbestand av mort og abbor, og med antatt dominans av karuss er derfor forventet fiskesamfunn dersom oksygensvinn er den begrensende faktor. Sekundæreffekter av dette er at fisk i perioder med dårlig oksygenkonsentrasjon vil søke til områder av innsjøen som fortsatt har tilfredsstillende forhold, noe som vil forventes å gi hard predasjon på småfisk og derved dårlig rekruttering.
Men flere forhold tilsier at oksygensvinn i vannmassene alene ikke er den begrensende faktoren i årene forut for prøvefisket i 1998. Høsten 1995 ble det for det første påvist massiv dødelighet hos årsunger av abbor uten at oksygensvinn ble påvist. For det andre antyder oksygenmålinger utført av OVA på ettervinteren at forholdene er blitt bedre de 6-8 siste år sammenliknet med tilsvarende målinger på 1970- og 1980-tallet. Observasjon av større forekomster av vanlig dammusling (Anodonta piscinalis) høsten 1998 med mange årsklasser tilstede viser for det tredje god overlevelse av disse organismene, og det har trolig også vært en markert økning i denne bestanden de siste 5 årene, idet bare få individer ble påvist i 1993.
Bruk av vanlig dammusling som en indikasjon på oksygenforholdene må imidlertid gjøres med forsiktighet, fordi tålegrensene for oksygen ikke er avklart når det gjelder vanlig dammusling. Oksygensvinn anses imidlertid som årsaken til nærmest fravær av vanlig dammusling over flere tiår, selv om også gift fra blågrønnalger er nevnt (Økland & Økland 1991). En annen muslingart, svanemuslingen (Anodonta cygnea), har en viss toleranse ovenfor oksygensvinn, og det er vist at denne muslingarten tåler 7 døgn i oksygenfritt miljø i romtemperatur (Salanki 1977). Det må derfor regnes med at også vanlig dammusling har en viss toleranse overfor oksygensvinn, men det konkluderes med at god overlevelse og økning i bestanden av vanlig dammusling har skjedd de siste år som en følge av bedre oksygenforhold. På den andre siden utelukker ikke forekomst av vanlig dammusling at oksygensvinn fortsatt skjer og at dette kan gi dødelighet for fisk.
Utover oksygensvinn som mulig dødelighetsfaktor bør andre faktorer vurderes, og giftproduserende blågrønnalger anses her som en mulighet. Selv om det ikke er dokumentert effekter av giftige blågrønnalger på fisk i Østensjøvannet, bør direkte dødelighet og/eller subletale effekter på fisk vurderes. NIVA har foretatt spredte observasjoner av forekomsten av blågrønnalger og toxinproduserende stammer i Østensjøvannet. De framtredende artene i perioden 1980-98 har vært innenfor slekteneAnabaena ogMicrocystis(Olav Skulberg, pers. medd.). Det er foretatt biotester for akutt toksisitet og kjemiske analyser som har vist at blågrønnalger i Østensjøvannet produserer cyanotoksiner med effekt på lever og at dette er gifter som har en viss langtidsvirkning.
Det er ikke kjent om dette har en virkning på fisk, men vannet er meget grunt og har svært liten gjennomstrømning. Dette gir ingen mulighet for fisk å rømme ned under det algeproduserende sjikt eller til innløpsos i kritiske perioder. Disse forholdene sammen med høye konsentrasjoner av blågrønnalger som periodevis utvikles i Østensjøvannet skulle tilsi maksimal effekt av en eventuell algegift. Hvilken økologisk konsekvens dette skulle ha på fiskesamfunnet vil imidlertid bare være spekulasjoner, men det er lett å tenke seg at ungstadier har stor følsomhet, og at mort som konsumerer alger, deriblant blågrønnalger (Lessmark 1983), vil kunne bli mer påvirket enn abbor, både gjennom direkte dødelighet og gjennom subletale effekter. Det kan også spekuleres på om subletale effekter kan gi stor variasjon i vekstmønster mellom individene i populasjonen, ved at noen årsklasser blir mer eksponert enn andre.
Den praktiske konsekvensen av dette vil være at de begrensende faktorer vil endre karakter etterhvert som vannkvaliteten eventuelt bedres. Fra å være begrenset av en faktor som periodevis gir dødelighet (oksygensvinn, algegift) eller dårlig vekst, forventes det at bestandene etter hvert vil bli næringsbegrenset. Dette innebærer følgende endringer:
Økt biomasse og antall av fisk vil innebære at den relative betydningen av fiskens bidrag til eutrofiering vil øke. Dette vil være prosesser som må karakteriseres som langtidsendringer og som vil gå parallelt med en eventuell bedring av vannkvaliteten.
Forholdene i innsjøen må uansett karakteriseres som ustabile, også på lang sikt. Ustabiliteten mht. fisk henger sammen med at perioder med oksygensvinn opplagt kan inntreffe som en følge av høy produksjon og naturgitte forhold som lite vannvolum og lav gjennomstrømning som følge av liten tilførsel fra det lokale nedbørfelt. Det henvises her til episoden med oksygensvinn i 1903 og 1909 (Huitfeldt-Kaas 1909). Gjennomstrømning og tilførsel av vann er trolig en nøkkelfaktor for å stabilisere forholdene, og økt gjennomstrømning kan gi økt overlevelse av fisk i kritiske perioder.